אינדקס
אינדקס

Severe Acute Respiratory Syndrome Coronavirus 2−Specific Antibody Responses in Coronavirus Disease 2019 Patients
https://wwwnc.cdc.gov/eid/article/26/7/20-0841_article
Abstract
A new coronavirus, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), has recently emerged to cause a human pandemic. Although molecular diagnostic tests were rapidly developed, serologic assays are still lacking, yet urgently needed. Validated serologic assays are needed for contact tracing, identifying the viral reservoir, and epidemiologic studies. We developed serologic assays for detection of SARS-CoV-2 neutralizing, spike protein–specific, and nucleocapsid-specific antibodies. Using serum samples from patients with PCR-confirmed SARS-CoV-2 infections, other coronaviruses, or other respiratory pathogenic infections, we validated and tested various antigens in different in-house and commercial ELISAs. We demonstrated that most PCR-confirmed SARS-CoV-2–infected persons seroconverted by 2 weeks after disease onset. We found that commercial S1 IgG or IgA ELISAs were of lower specificity, and sensitivity varied between the 2 assays; the IgA ELISA showed higher sensitivity. Overall, the validated assays described can be instrumental for detection of SARS-CoV-2–specific antibodies for diagnostic, seroepidemiologic, and vaccine evaluation studies.In December 2019, a new coronavirus emerged in China and caused an acute respiratory disease now known as coronavirus disease 2019 (COVID-19) (1). The virus was identified to be a betacoronavirus related to severe acute respiratory syndrome coronavirus (SARS-CoV) and thus was named SARS-CoV-2 (2). In <2 decades, this virus is the third known coronavirus to cross the species barrier and cause severe respiratory infections in humans after SARS-CoV in 2003 and Middle East respiratory syndrome coronavirus (MERS-CoV) in 2012, yet with unprecedented spread compared with the earlier 2 viruses.
Because of the rapid increase in number of cases and uncontrolled and vast spread worldwide, the World Health Organization has declared SARS-CoV-2 a pandemic. As of March 14, 2020, the virus had infected >130,000 persons in 122 countries, 3.7% of whom had died. (3). Rapid identification of the etiology and sharing of the genetic sequence of the virus, followed by international collaborative efforts initiated because of emergence of SARS-CoV-2, has led to rapid availability of real-time PCR diagnostic assays that support case ascertainment and tracking of the outbreak (4). Availability of these assays has helped in patient detection and efforts to contain the virus. However, validated serologic assays are still lacking and are urgently needed.
Validated serologic assays are crucial for patient contact tracing, identifying the viral reservoir hosts, and epidemiologic studies. Epidemiologic studies are urgently needed to help uncover the burden of disease, in particular the rate of asymptomatic infections, and to get better estimates on illness and death. In addition, these epidemiologic studies can help identify the extent of virus spread in households, communities, and specific settings, which could help guide control measures Serologic assays are also needed for evaluation of results of vaccine trials and development of therapeutic antibodies.
Among the 4 coronavirus structural proteins, the spike (S) and nucleocapsid (N) proteins are the main immunogens (5). We describe development of serologic assays for detection of virus neutralizing antibodies and antibodies to the N protein and various S protein domains, including the S1 subunit, and the receptor-binding domain (RBD) of SARS-CoV-2 in an ELISA format. Using a well-characterized cohort of serum samples from PCR-confirmed SARS-CoV-2 and patients PCR-confirmed to be infected with seasonal coronaviruses and other respiratory pathogens, we validated and tested various antigens in different platforms developed in-house, as well as a commercial platform.
[size=21]Materials and Methods
Serum Samples
Erasmus Medical Center Samples
We used serum samples (n = 10) collected from 3 PCR-confirmed patients: 2 with mild COVID-19 and 1 with severe COVID-19 (Table 1) from France in accordance with local ethics approvals (F.-X. Lescure et al., unpub. data, https://doi.org/10.1101/2020.03.11.987958External Link). For assay validation, we used samples obtained from persons who had PCR-diagnosed infections with human coronaviruses (HCoV-229E, NL63, or OC43), SARS-CoV, MERS-CoV, or other respiratory viruses (Table 1) as reported (6). We also included samples from patients who had recent infections with cytomegalovirus, Epstein-Barr virus, or Mycoplasma pneumoniae because these pathogens have a higher likelihood of causing false-positive results. As negative controls, we used serum samples from 45 healthy blood donors (Sanquin Blood Bank, https://www.sanquin.nlExternal Link) (cohort A). We also tested serum samples from SARS patients (7). All samples were stored at −20°C until use. The Sanquin Blood Bank obtained written informed consent for research use of samples from blood donors. Use of serum samples from the Netherlands was approved by the local medical ethics committee (approval no. 2014–414).
Berlin Samples
All serum samples (n = 31) from patients with PCR-confirmed cases of COVID-19 cases were previously analyzed by a recombinant SARS-CoV-2 S protein–based immunofluorescence test and plaque reduction neutralization (R. Wölfel et al., unpub. data, https://doi.org/10.1101/2020.03.05.20030502External Link). We tested serum samples as part of an extended diagnostic regimen after we obtained informed written consent from patients. We obtained non–SARS-CoV-2–infected serum samples (n = 31) from the serum collection of the National Consiliary Laboratory for Coronavirus Detection at Charité–Universitätsmedizin Berlin (Berlin, Germany). Samples were collected after we obtained informed written consent. The collection contained follow-up antibody-positive serum samples from PCR-confirmed virus-infected cases: HCoV-229E (n = 4), HCoV-HKU1 (n = 3), HCoV-OC43 (n = 7), MERS-CoV (n = 3), HCoV-NL63 (n = 6), SARS-CoV (n = 3), and common cold CoV (n = 6).
Protein Expression
We expressed the S ectodomains of SARS-CoV-2 (residues 1–1,213, strain Wuhan-Hu-1, GenBank accession no. QHD43416.1), SARS-CoV (residues 1–1,182, strain CUHK-W1, accession no. AAP13567.1), and MERS-CoV (residues 1–1262, strain EMC, accession no. YP_009047204.1) in HEK-293T cells by using a C-terminal trimerization motif, Strep-tag, and the pCAGGS expression plasmid. Likewise, we expressed the SARS-CoV-2 S1 subunit or its subdomains (S;S1, residues 1–682; S1A, residues 1–294; RBD, residues 329–538; accession no. QHD43416.1) in 293T cells, as described (C. Wang et al., unpub. data, https://doi.org/10.1101/2020.03.11.987958External Link).
We produced S1 proteins of other HCoVs: HKU1 (residues 1–750), OC43 (residues 1–760), NL63 (residues 1–717), 229E (residues 1–537), SARS-CoV (residues 1–676), and MERS-CoV as described (6,8). We affinity purified all recombinant proteins from culture supernatant by using Protein-A Sepharose beads (catalog no. 17–0780–01; GE Healthcare, GE Healthcare, https://www.gehealthcare.comExternal Link) or strep-tactin beads (catalog no. 2–1201–010; IBA Lifesciences, https://www.iba-lifesciences.comExternal Link). We checked purity and integrity of all purified recombinant proteins by using sodium dodecyl sulfate–polyacrylamide gel electrophoresis and staining with Coomassie blue.
Plaque Reduction Neutralization Test
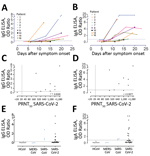
We used the plaque reduction neutralization test (PRNT) as a reference for this study because neutralization assays are the standard for coronavirus serologic analysis. We tested serum samples for their neutralization capacity against SARS-CoV-2 (German isolate; GISAID ID EPI_ISL 406862; European Virus Archive Global #026V-03883) by using PRNT as described with some modifications (9). We 2-fold serially diluted heat-inactivated samples in Dulbecco modified Eagle medium supplemented with NaHCO3, HEPES buffer, penicillin, streptomycin, and 1% fetal bovine serum, starting at a dilution of 1:10 in 50 μL. We then added 50 μL of virus suspension (400 plaque-forming units) to each well and incubated at 37°C for 1 h before placing the mixtures on Vero-E6 cells. After incubation for 1 h, we washed, cells supplemented with medium, and incubated for 8 h. After incubation, we fixed the cells with 4% formaldehyde/phosphate-buffered saline (PBS) and stained the cells with polyclonal rabbit anti-SARS-CoV antibody (Sino Biological, https://www.sinobiological.comExternal Link) and a secondary peroxidase-labeled goat anti-rabbit IgG (Dako, https://www.agilent.comExternal Link). We developed signal by using a precipitate forming 3,3′,5,5′-tetramethylbenzidine substrate (True Blue; Kirkegaard and Perry Laboratories, https://www.seracare.comExternal Link) and counted the number of infected cells per well by using an ImmunoSpot Image Analyzer (CTL Europe GmbH, https://www.immunospot.euExternal Link). The serum neutralization titer is the reciprocal of the highest dilution resulting in an infection reduction of >50% (PRNT50). We considered a titer >20 to be positive.
We performed the PRNT for serum samples from Germany by using Vero E6 cells, as described (R. Wölfel et al., unpub. data, https://doi.org/10.1101/2020.03.05.20030502External Link) (10) and 24-well plates. Before the PRNT, we heat-inactivated patient serum samples at 56°C for 30 min. For each dilution step (in duplicate), we diluted patient serum samples in 200 μL of OptiPro serum-free medium (https://www.thermofisher.comExternal Link) and mixed 1:1 with 200 μL of virus solution containing 100 PFUs. We vortexed the 400-μL serum–virus solution gently, incubated at 37°C for 1 h, and then incubated each 24-well plate with 200 μL serum–virus solution. After incubation for 1 h at 37°C, we discarded supernatants, washed cells once with PBS, and supplemented them with 1.2% microcrystalline cellulose solution in Dulbecco modified Eagle medium. After 3 days, we fixed and inactivated the plates by using a 6% formaldehyde/PBS solution and stained with crystal violet.
ELISA
We performed anti-SARS-CoV-2 S1 IgG and IgA ELISAs by using β-versions of 2 commercial kits (EUROIMMUN Medizinische Labordiagnostika AG, https://www.euroimmun.comExternal Link) and performed the assay according to the manufacturer’s protocol. We detected optical density (OD) at 450 nm and calculated a ratio of the reading of each sample to the reading of the calibrator, included in the kit, for each sample (OD ratio). Because the β-version of the kit awaits validation and marking, we determined an in-house cutoff value based on the mean background reactivity of all SARS-CoV-2–negative serum samples in the study multiplied by 3. The OD ratio was 0.9 for IgA and 0.3 for IgG.
We performed the in-house ELISAs by coating 96-well microtiter ELISA plates with in-house–produced S antigens (S or S1 of SARS-CoV-2, SARS-CoV or MERS-CoV; SARS-CoV-2 S1A; or RBD proteins) or SARS-CoV N protein (Sino Biological) in PBS overnight at 4°C. After blocking, we added diluted serum (diluted 1:100 or 2-fold serially diluted for titers) and incubated at 37°C for 1 h. Antigen-specific antibodies were detected by using peroxidase-labeled rabbit anti-human IgG (Dako) and 3,3′,5,5′-tetramethylbenzidine as a substrate. The absorbance of each sample was measured at 450 nm, and we set the cutoff value at 6 SD above the mean value for the negative cohort.
S1 Protein Microarray
Serum samples were previously tested for antibodies against S1 of different coronaviruses. We used a protein microassay that has been described (6).
Statistical Analysis
We analyzed the correlations between antibody responses detected by different ELISAs and those detected by PRNT, which is the standard for coronavirus serologic analysis. We used GraphPad Prism version 8 (https://www.graphpad.comExternal Link) for this analysis.
[/size]